
Hey there! If you’re an A-Level Biology student looking for help with eukaryotic organelles, then you’re in the right place! This is a pretty important topic as it will also lay the foundation for the rest of the syllabus over your 2 years in JC.
For example, the topic Genetics and Inheritance will focus a lot on the nucleus and its components, Respiration and Photosynthesis will explore the mitochondria and chloroplast in detail, and Cell Signalling would look at how a signal transverses a cell from its exterior environment, to the nucleus and all the way back on itself.
“It’s so much information to memorise!” Yes! It is a lot, but here’s a secret: The examiners are looking for keywords that demonstrate your understanding of the question.
What are these magic words, and how do I know when to use them?
In this article, I have summarized the key concepts on cellular organelles that are most commonly tested and will make frequent appearances in future chapters, along with keywords to help communicate your understanding to your examiner.
While this article is just a brief overview of a vast subject, I hope you stay humbly curious, and use this resource to kick start your journey. With that, let us begin this article!
The human body is made of trillions of eukaryotic cells of various cell types, each working hard to keep you alive. Each cell is hailed as a microscopic building block of life, measuring on average 10 to 100 micrometers in diameter. But, imagine an entire functional city within your cells, with each building performing a specialized function to keep the cell running. These “buildings” are the organelles within the eukaryotic cell. And just like the walls of a building, each organelle is membrane bound and part of a sophisticated endomembrane system, allowing it to maintain its own optimal conditions to carry out its function.
On the other hand, prokaryotes are much simpler creatures. Unlike eukaryotes, prokaryotes generally lack membrane-bound organelles. The endosymbiotic theory suggests that organelles like the mitochondria and chloroplast descend from ancient prokaryotes that were swallowed by a larger cell (the endo- part of the theory). These prokaryotes co-existed with their hosts, eventually evolving into organelles (the -symbiotic part).
In this article, we will go on a tour around the eukaryotic organelles, their functions and how to tell them apart in those grainy transmission electron microscope (TEM) images.
But before you go on reading… You might want to download a pdf copy of this article as it is quite long!
Click the ‘Download’ button, enter your email, and the pdf file will be delivered to your inbox! (Remember to check spam!)

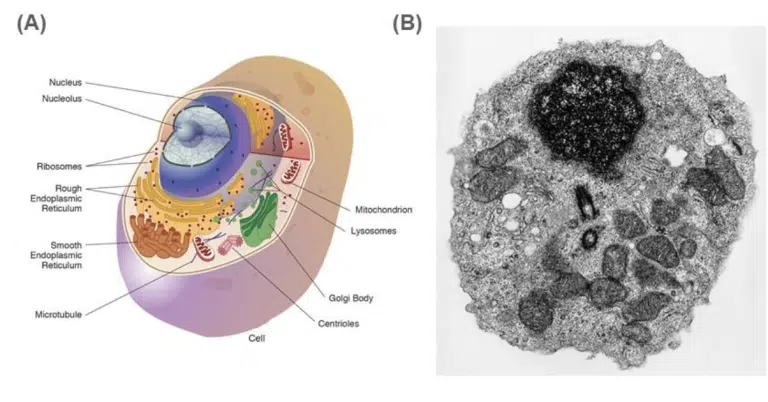
Fig 1. (A) Illustration and (B) Transmission electron microscopy image of an eukaryotic cell and its organelles.
The nucleus stores and protects the cell’s DNA, and is found in all eukaryotic cells except for mature red blood cells. It is usually spherical or ovoid, and 10 to 20 micrometers in diameter. DNA replication, transcription and gene expression occurs here. Some cell types (eg. skeletal muscle cells) may have more than one nucleus!
The nucleolus appears as a large and densely stained area within the nucleus. More than one nucleoli may be present in each nucleus. The nucleolus produces ribosomal RNA (rRNA) through transcription of rRNA-encoding DNA. These rRNA molecules are assembled with ribosomal proteins imported from the cytosol to form the small and large ribosomal subunits.
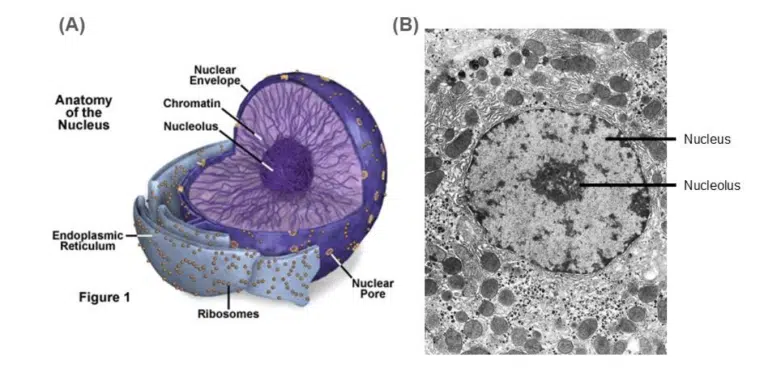
Fig 2. (A) Illustration and (B) Transmission electron microscopy image of the nuclei and the surrounding organelles.
The nucleus is a spherical structure. Regions of highly condensed DNA (heterochromatin) can be seen as darker spots, while regions of loosely packed DNA (euchromatin) can be seen as lighter spots. At certain stages of the cell cycle (especially when rRNA is in high demand), the nucleolus can be seen as a dense spot within the nucleus.
Ribosomes comprise of rRNA and ribosomal proteins and are synthesized in the nucleolus. Despite lacking a membrane, ribosomes are still considered organelles as they have the distinct function of producing proteins. Each ribosome is made of a small and large subunit. During translation, the small and large subunits assemble in the cytosol to form a functional ribosome. 70S ribosomes can be found in prokaryotes, mitochondria and chloroplasts (remember the endosymbiotic theory!) 80S ribosomes are found in eukaryotes, and can exist either as free ribosomes in the cytosol or fixed ribosomes on the rough endoplasmic reticulum.
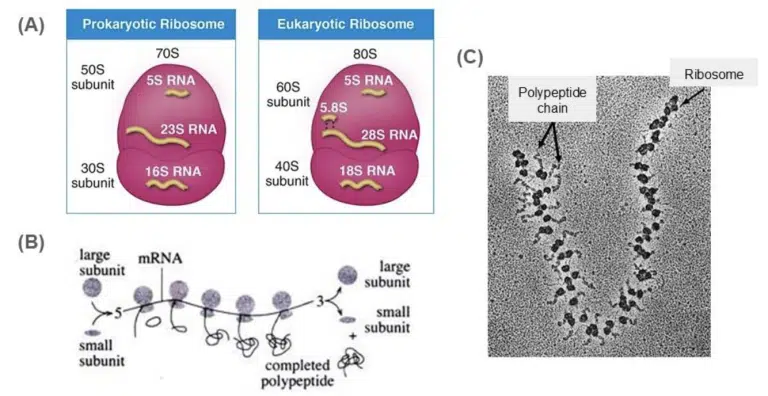
Fig 3. (A) Illustration of the 70S prokaryotic and 80S prokaryotic ribosome. (B) Illustration of a polysome, which refers to a group of ribosomes bound to and translating a single mRNA strand. (C)Transmission electron microscope image of a polysome. The growing polypeptide chain can be observed extending from each ribosome bound to the mRNA.
The endoplasmic reticulum (ER) is a network of membranes within the cell that is continuous with the nuclear membrane. Based on the shape of the lumen and the presence of ribosomes, we can classify the ER into the rough ER and the smooth ER.
The rough ER is characterised with flattened cisternae (sacs) and the presence of bound ribosomes. Ribosomes producing proteins meant to be secreted dock here, resulting in a “rough” spotty appearance. Secretory proteins are produced here.
The smooth ER is smooth… as it is not associated with ribosomes. It consists of tubular cisternae and has a variety of functions (lipid synthesis, metabolism of carbohydrates, detoxification of drugs in the liver and storage of Ca2+ ions).
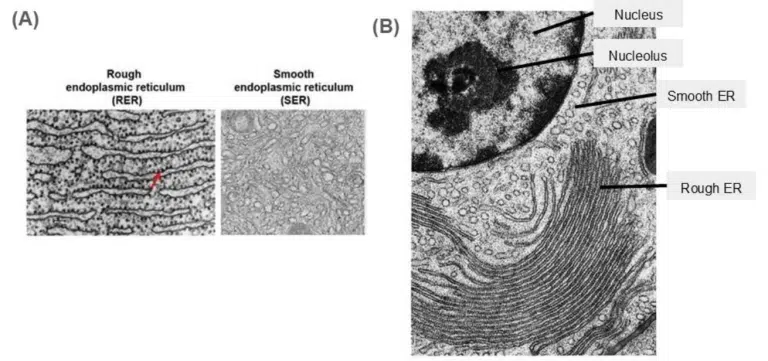
Fig 4. (A) The difference between the rough ER and the smooth ER as seen under transmission electron microscopy. The rough ER is characterized by its “rough” dotted appearance due to the presence of bound ribosomes, and its flattened cisternae. The smooth ER lacks these bound ribosomes and appear as tubular structures in contrast to the rough ER’s flattened appearance. (B) The transmission electron microscope image of a nucleus, smooth ER and rough ER. A quick way to find the ER endomembrane system is to look near the nucleus! Their membranes are connected after all, so the ER would be located right next to the nucleus. Observe the difference in shape between the smooth ER and rough ER in this image!
The Golgi is made up of a stack of flattened cisternae. This is where proteins produced at the rough ER undergo further biochemical modifications. The cis face of the Golgi faces the nucleus and ER, and hence is the receiving end of incoming protein-loaded vesicles. Products from the ER move across the Golgi from the cis face to the trans face via repeated budding and fusing of vesicles. These protein products are packaged into vesicles at the trans face and bud off to their final destinations. Depending on their content, these vesicles could be:
In plant cells, Golgi-derived vesicles carry cell wall materials and form the cell plate during cellular division.
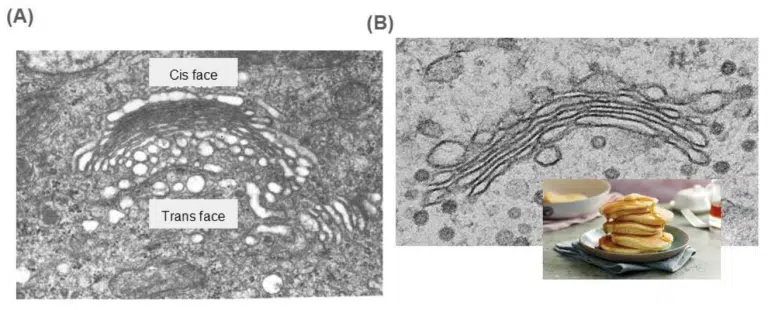
Fig 5. (A) A transmission electron microscope image of the Golgi. While not always obvious, the Golgi tends to curve slightly towards the trans face. A way to identify the trans face is to look for vesicles budding off! The trans face also faces away from the nucleus and ER, towards the exterior of the cell as protein products exit the Golgi and head towards their final destination. (B) The Golgi resembles a stack of pancakes. Unlike the rough ER where the cisternae is a continuous endomembrane system, the Golgi cisternae are separate from each other.
Lysosomes can be considered the clean-up crew of the cell. They contain hydrolytic enzymes and are membrane-bound to prevent its contents from damaging the cell. The internal environment of the lysosome is acidic, as its enzymatic contents require an optimal pH of 5. The functions of lysosomes include intracellular digestion, autophagy (digestion of damaged cellular components) and autolysis (self-destruction of the cell). Lysosomes appear as spherical sacs 2 to 5 um in diameter. Lysosomes can be considered either:
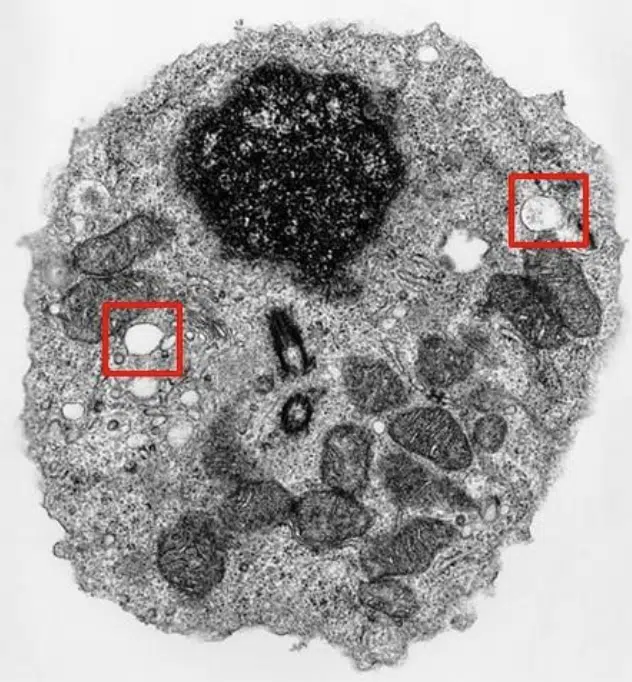
Fig 6. Transmission electron microscopy image of a cell, with 2 lysosomes highlighted.
Lysosomes are vesicles filled with hydrolytic enzymes, and maintain a low internal pH to facilitate the function of its enzymes. Lysosomes are found across the cytosol of the cell.
The powerhouse of the cell! The mitochondria is characterized by a double membrane structure, where the inner membrane folds extensively to form wrinkles known as cristae. ATP is produced here as a product of cellular respiration. As an endosymbiotic organelle, the mitochondria contains its own circular mitochondrial DNA (mtDNA) and 70S ribosomes.
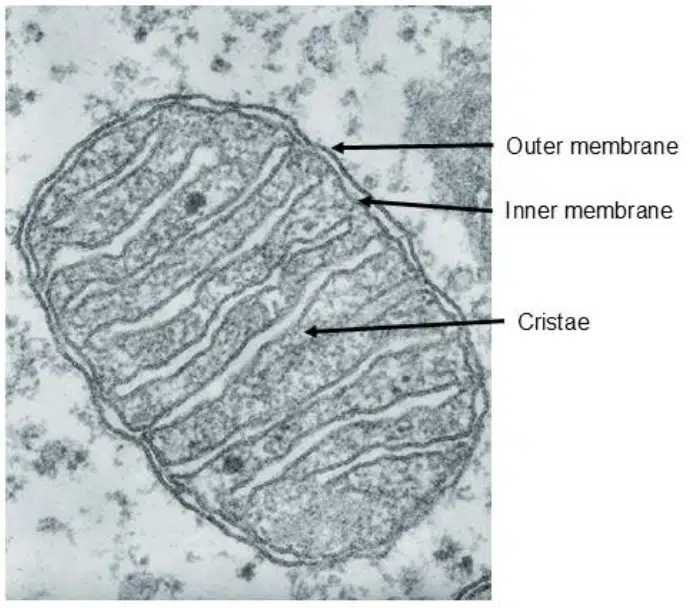
Fig 7. Transmission electron microscopy image of a mitochondria.
The mitochondria is a double membrane system consisting of the outer and inner membrane, with an intermembrane space in between. The inner membrane folds to form cristae, which are the most recognizable feature of mitochondria. The cristae increase the surface area of the inner membrane, allowing space for more enzymes involved in cellular respiration to produce more ATP.
Microtubules can be considered the structural support of the cell. They are long polymers of the protein tubulin and form part of the cell’s cytoskeleton. Centrioles are small barrel-shaped organelles found within a region known as the centrosome, and exist in most eukaryotes except in higher plants. Centrioles are found in pairs positioned 90° from each other. The centrosome is usually located near the nucleus, except during cell division where it replicates and moves towards the poles of the cell. The centrosome is also regarded as the microtubule organising center (MTOC), and functions to organize microtubules during cellular division.
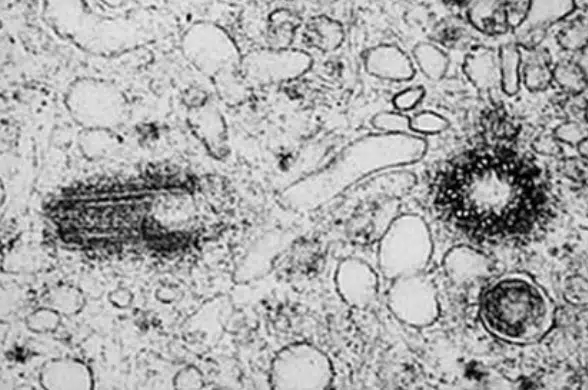
Fig 8. Transmission electron microscopy image of the centrioles. They occur in pairs, where one barrel is rotated 90° from the other.
The membrane that separates and protects the cell from its external environment. The plasma membrane is composed of lipids (mostly phospholipids), proteins and carbohydrates. The phospholipids are arranged as a bilayer, where the hydrophilic heads face the aqueous nature of the cell and the external environment and the hydrophobic tails form the core of the plasma membrane.
The plasma membrane structure is commonly described using the fluid mosaic model, where the flexible, fluid phospholipid bilayers are embedded asymmetrically with various proteins (the mosaic). The components of the plasma membrane move freely laterally within the membrane, but not across the membrane.
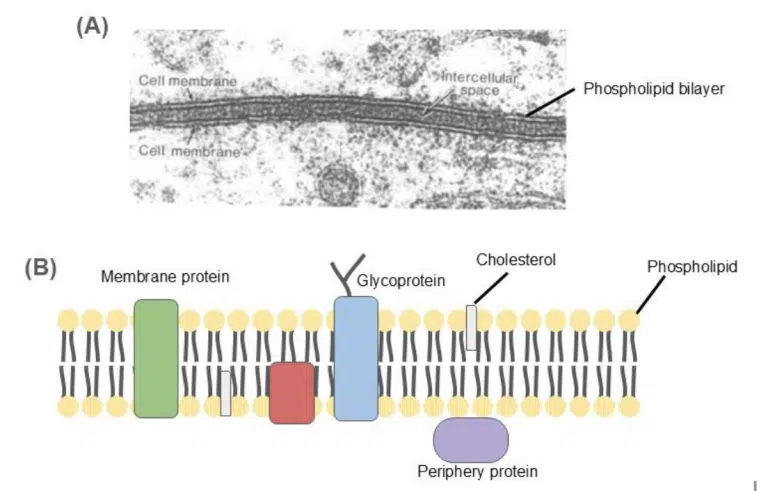
Fig 9. (A) Transmission electron microscopy image of the cell membranes of 2 adjacent cells. The bilayer structure of each plasma membrane can be seen. (B) A schematic illustration of the fluid mosaic model. The plasma membrane is made of the phospholipid bilayer, with proteins and cholesterol dotted across the membrane. The components of the plasma membrane can move freely within its lateral confines, but flipping across the membrane is highly unlikely.
Now that we have journeyed across the organelles making up a eukaryotic animal cell, let’s take a look at some extra organelles that are present in the eukaryotic plant cell.
The chloroplast is the site of photosynthesis and produces oxygen from carbon dioxide. It is characterized by its triple membrane structure and coin-like stacks of thylakoids (granum, plural grana) inside its stroma. Like the mitochondria, the chloroplast contains its own circular DNA genome known as chloroplast DNA (cpDNA) and 70S ribosomes.
The plant lacks a skeletal system, and thus needs another form of structural support to keep it standing! The plant cell wall is a rigid, semi-permeable barrier that surrounds plant cells. It is made primarily from cellulose and lignin. The plant cell wall protects the cell from mechanical stressors, provides rigid shape and support to the cell. To facilitate movement of materials between adjacent plant cells, channels called plasmodesmata can be found between the cell walls.
The central vacuole is a large fluid-filled organelle in the plant cell. Its main purpose is to maintain turbid pressure within the plant cell much like how air keeps a tire inflated.
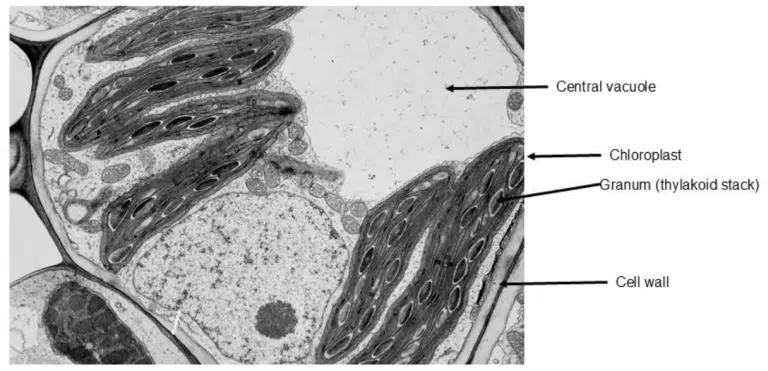
Fig 10. Transmission electron microscopy image of a plant cell.
Some unique organelles from the plant cell are labelled. The central vacuole can be identified as a large space within the cell. Chloroplasts can be recognised by the thylakoid stacks (grana) within its structure. Compared to mitochondria, chloroplasts also appear flatter and more disc-like. The cell wall is the thick layer of cellulose that surrounds the plant cell, and can been seen as the wide margin that encloses the plant cell.
By following a protein’s synthesis in a cell, we can observe how various organelles of the eukaryotic cell function together as a whole. Protein synthesis begins in the nucleus, where the encoding gene is transcribed into mRNA. The mRNA is transported to the cytosol, where ribosomes bind to the mRNA and initiate its translation into a polypeptide. Depending on whether the protein is designated for transport to the Golgi and subsequent export, the protein may encode a signal peptide.
If the protein is designated to be secreted, a signal peptide will be encoded and the protein would undergo the secretory pathway. Upon translation of this signal peptide, the ribosome would associate itself with the rough ER, where translation of the polypeptide continues inside the lumen of the ER. Here, the growing polypeptide chain folds into its 3D conformation and undergoes some biochemical modification, such as glycosylation. The folded protein is then transported from the rough ER to the cis face of the Golgi. Once at the Golgi, the protein transits from the cis to trans face of the Golgi as further modifications are made. Such modifications may include glycosylation and hydroxylation. At the trans face, the modified protein would be sorted and packaged into secretory vesicles which bud off the Golgi to be transported to various cellular destinations. These vesicles would end up:
For proteins that are not intended for export, there would not be a signal peptide encoded and the protein would be synthesized by a free ribosome in the cytosol.
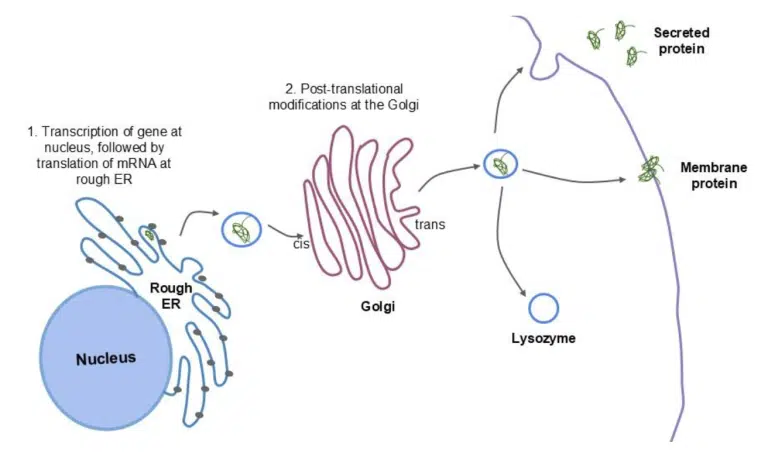
Fig 11. Schematic diagram of the secretory pathway.
The gene encoding the protein is transcribed into mRNA at the nucleus. The mRNA is translated into protein by bound ribosomes at the rough ER, where the protein is folded into its 3D conformation within the ER lumen. The protein is transported to the cis face of the Golgi, and undergoes further modifications as it transits towards the trans face. The completed proteins are sorted and packaged into vesicles that bud off the trans face to be transported to their final destination.
In conclusion, organelles play specialized and crucial roles in ensuring the survival, growth and function of the eukaryotic cell. This article briefly covers the functions of key organelles, and how they work together such as in the secretory pathway. In future chapters, we will have the opportunity to delve deeper into some of these organelles (the nucleus in genetics, centrioles in cellular division, mitochondria in oxidative respiration, chloroplast in photosynthesis etc.). Until then, do make sure that you are familiar with the basics of each organelle! It’ll get easier from here on!
You might want to download a pdf copy of this article for future reference!
Click the white download button below, enter your email, and the pdf file will be delivered to your inbox! (Remember to check spam!)
The Science of Studying provides live online tuition via Zoom classes for Combined/Pure Chemistry, Biology, and Physics. To date, we have taught 800+ students over 12 years.
In case you are wondering, yes – there is a science behind studying!
At Science of Studying, we use our SOS system™ to teach our classes so that even last-minute students can see remarkable improvements in their grades – without mind-numbing memorisation of textbooks and without the drudgery of doing numerous assessment books.
All these conducted in a fun, interactive, stress-free online environment.
If you need help with your Chemistry, Biology, and Physics subjects, do reach out to us and we will see what we can do to help.
Contact Us: Click Here
Admin number: +65 88082348

The SOS system™️ guides students through an effective process of:
Join our proven online tuition programs and see real improvements in understanding, confidence, and school results.
Book a free trial lesson and start the journey today or discover more below:
WhatsApp us